The Guide-it Knockin Screening Kit provides a fast and accurate assay for detecting the successful introduction of SNPs, both in mixed populations and in clonal cell lines
The dual-color technology employed by the kit allows simultaneous detection of two alleles, enabling identification of heterozygous cell lines carrying one copy each of SNP and WT alleles
The ease with which somatic cells obtained from patient and nonpatient populations can be reprogrammed into human induced pluripotent stem cells (iPSCs) and edited in a precise manner (using CRISPR or related technologies) has dramatically expanded the applicability and accessibility of iPSC-derived cell lines for disease modeling and drug development (Figure 1). Introduction of disease-associated genetic variants, including SNPs, in iPSCs allows for the evaluation of the phenotypic effects of these variants in relevant cell types and eliminates variability due to genetic background, thus enabling finer analysis of disease mechanisms and more rapid assessment of potential therapies (e.g., drug screening). Looking a step further, a promising clinical application for precise genome editing could be the correction of pathogenic mutations in patient-derived iPSCs and delivery of the engineered cells to the patients as therapy.
Figure 1. Applications for genome editing in iPSCs. The genetic modification of iPSC lines to introduce pathogenic mutations or correct them has enabled the generation of isogenic cell lines for disease modeling and drug screening applications. In the future, the correction of disease-associated mutations in patient-derived iPSCs could also provide a basis for novel therapeutic approaches. Image from Kim, Kang & Ju, 2017 used under creative commons license. Copyright @ 2017 The Korean Association of Internal Medicine.
Workflow
One of the most common strategies in creating disease models is the use of CRISPR/Cas9 in tandem with single-stranded donor oligonucleotide (ssODN) encoding the desired modification, but this approach relies on homology-directed repair (HDR), an inefficient endogenous repair mechanism that typically results in a low percentage of successfully edited cells.
Genome editing experiments involving the application of HDR to generate SNPs typically yield three possible outcomes at the target site: retention of the wild-type (WT) sequence (i.e., no editing), creation of an insertion or deletion (Indel), or successful HDR (SNP) (Figure 2, Panel A). These outcomes can be distributed in varying allelic combinations in diploid cells (Figure 2, Panel B). When studying diseases caused by dominant alleles, researchers often want to generate isogenic cell lines with one allele edited (encoding the SNP of interest) while maintaining the other allele as wild type. This outcome is particularly difficult to achieve since most cells edited via HDR are either homozygous for the SNP (SNP/SNP) or carry an indel allele in addition to the SNP allele (SNP/Indel). This challenge, together with the low efficiency of HDR, has necessitated the optimization of the editing conditions (to maximize HDR efficiency) and the development of fast and reliable methods for screening large numbers of clonal cell lines.
Figure 2. Common outcomes when engineering SNPs. An example of a single-nucleotide edit (G>T) is shown. Panel A. Outcomes at the genomic target site. When cleavage fails to occur at the target site or is followed by accurate, nonhomologous end joining (NHEJ)-based repair, the result is the wild-type (WT) sequence. When cleavage is followed by inaccurate NHEJ-based repair, the result is an insertion or deletion (Indel) at the target site, possibly causing a knockout (KO; a highly probable outcome). When cleavage is followed by accurate HDR, a SNP is introduced at the target site. Panel B. Combined allelic outcomes in diploid cells. When editing is performed in diploid cells, the outcomes for each allele can vary, generating multiple possible combinations. Cells can remain homozygous (Wild type; top), they can have one or both alleles modified via inaccurate NHEJ (Indel; middle), or they can have one or both alleles modified with the desired SNP (Successful HDR; bottom).
The Guide-it Knockin Screening Kit offers the possibility to directly analyze HDR efficiency in bulk-edited populations, allowing researchers to move forward with isolation of single-cell clones from populations that have the highest frequency of successful HDR. Subsequently, clonal cell lines can be screened simultaneously for two specific edits (e.g., SNP/WT).
The first step of the workflow for the Guide-it Knockin Screening Kit involves PCR amplification of the targeted genomic locus. The resulting PCR product is then hybridized with two user-designed oligos encoding the sequence to be detected. If there is full hybridization between the oligos and the PCR product, an enzymatic reaction that generates a fluorescent signal is triggered, such that detection of a fluorescent signal is correlated with the presence of a specific sequence at the target site. The assay is sensitive enough to detect single-base differences, and the dual fluorescence capability (red/green) of the kit allows for the simultaneous detection of two different sequences at the same target site (Figure 3). Therefore, the kit can be used to screen for heterozygous clones with one probe encoding the SNP allele (triggering a green signal) and another probe encoding the wild-type allele (generating a red signal).
Figure 3. SNP analysis workflow for the Guide-it Knockin Screening Kit. This example workflow demonstrates analysis of a G>A substitution, where G is the wild-type base edited to an A. After genome editing, single cells expanded to clonal cell lines can have several different genotypic outcomes at the genomic target site of interest (as shown in Figure 2, Panel B). After PCR amplification of the target site, the PCR product is annealed simultaneously with different oligo probes: a displacement oligo (purple) in combination with either flap-probe oligo A (green; encoding the SNP allele, A) or flap-probe oligo B (orange; encoding the WT allele, G). After the annealing of the oligos to the PCR products, the Guide-it Flapase enzyme (indicated with scissors) recognizes a complete base pairing and cleaves the 5′ portion of the flap-probe oligo (shaded green or orange). The cleaved flaps are then detected by corresponding Guide-it flap detectors, which yield green or red fluorescent signals, respectively. In the example above, analysis of a clonal cell line that is homozygous WT (G/G) at the site of interest yields only a red signal, while analysis of a heterozygous clone carrying both edited and WT alleles (G/A) yields both red and green signals.
In-house data
Detection of precise editing at a specific genomic locus in bulk-edited and clonal cell populations
One of the strategies to maximize the percentage of heterozygous clones (WT/SNP) is to use an equimolar mixture of ssODNs encoding the SNP or WT alleles and silent mutations in the PAM site (if possible) as HDR templates (Paquet et al. 2016). Mutation of the PAM site prevents Cas9 from recutting and generating indels at the genomic target site following successful HDR with either template, indirectly increasing the likelihood of obtaining heterozygotes.
We applied this method to generate an iPSC line heterozygous for a C>G substitution (S38C) in an endogenous gene of interest (unidentified for reasons of confidentiality). We electroporated two HDR templates (Figure 4, Panel A) encoding either the SNP (in blue) or the WT allele (in purple). The introduction of the SNP results in the destruction of the PAM site (CCG>GCG), and the oligo encoding the WT allele also codes for a silent mutation in the PAM sequence (C>G, in red; CCG>CGG).
The Guide-it Knockin Screening Kit was used to detect successful HDR of both templates (SNP or WT silent) in bulk-edited populations (Figure 4, Panel B). The lack of green and red fluorescent signals for cells electroporated with Cas9 protein in the absence of sgRNA ("negative control") or HDR templates ("KO") confirmed the specificity of the flap-probe oligos since they did not yield false-positive fluorescent signals in the presence of the unedited wild-type sequence or indel alleles.
Following editing and clonal cell isolation and expansion, the Guide-it Knockin Screening Kit was also used to identify clones with successful editing. Clones carrying the SNP or WT silent alleles could be detected with green or red fluorescence, respectively, but no heterozygotes carrying both edits (generating both red and green fluorescence signals; Figure 4, Panel C) were identified in this experiment, which had a low percentage of HDR.
Figure 4. Detection of precise editing at an endogenous locus in bulk-edited and clonal iPSC populations.Panel A. Editing outcomes following successful HDR at an anonymous locus of interest. Following successful HDR, the edited locus will encode either a SNP (in blue, lowercase) or a WT base (in purple) combined with a silent PAM mutation (in red, lowercase). Panel B. Detection of successful HDR in bulk-edited iPSCs. Displacement and flap-probe oligos were designed to detect WT silent or SNP alleles, yielding red and green fluorescent signals, respectively. In independent experiments, cells were electroporated with Cas9 protein alone (negative control), Cas9-sgRNA RNP complexes (KO), or RNP complexes combined with antisense SNP or SNP/WT silent ssODN mixtures. Synthetic oligos encoding the WT silent or SNP sequences were assayed in parallel as positive controls. For each editing scenario in which ssODNs were included in the electroporation mixture, successful HDR could be detected in the bulk population using the Guide-it Knockin Screening Kit, as indicated by the resulting fluorescent signals. Panel C. Detection of successful HDR in clonal cell lines. Clones obtained from single cells isolated by flow cytometry were screened for both edits (SNP and WT silent). While successful incorporation of either edit could be detected in separate clonal cell lines, no heterozygous clones carrying both edits were identified.
Comparing the results of the knockin screening kit with direct Sanger sequencing
The results of the Guide-it knockin screening were confirmed by Sanger sequencing. In cases where direct Sanger sequencing of the PCR reaction yielded ambiguous results or traces that were challenging to interpret (e.g., for instances where one of the edited alleles contained an indel), the genomic target region was amplified by PCR and cloned into pUC19 following the protocol of the Guide-it Indel Identification Kit. Sequencing of several bacterial clones in each case confirmed the results from the Guide-it Knockin Screening Kit assay.
Comparing the results of the knockin screening kit with direct Sanger sequencing demonstrated the challenges associated with interpreting data from the latter approach, even when the Sanger data was analyzed with the aid of a commonly used bioinformatics tool. To demonstrate this, we compared results obtained using either the knockin screening kit or Sanger sequencing to analyze a clonal cell line edited at an unidentified locus as described above (Figure 5). The knockin screening kit detected the presence of at least one correctly edited WT allele (WT silent), indicating that there could be two copies of the WT silent allele or that it could be paired with an allele encoding the unedited wild-type sequence or an indel. In contrast, direct Sanger sequencing of the PCR product amplified from the clonal cell line yielded a sequencing chromatogram with overlapping peaks at the region of interest (Figure 5, first chromatogram). Analysis of this sequencing data with a commonly used bioinformatic tool indicated that different alleles could be present in the sample at various probabilities (WT, 44%; WT silent, 17%; 1-bp deletion, 14%; another 1-bp deletion, 11%). Using the Guide-it Indel Identification Kit, it was determined that the clone had one correctly edited WT allele (WT silent; Figure 5, middle chromatogram) and one indel allele with a one-base insertion (G) together with a 7-bp deletion (Figure 5, bottom chromatogram). This result was consistent with the output of the knockin screening kit and demonstrated that the results obtained via bioinformatic analysis of Sanger sequencing could be misleading.
Figure 5. Comparing results from the Guide-it Knockin Screening Kit vs.Sanger sequencing. The knockin screening assay detected the presence of at least one correctly edited WT allele (WT silent; top sequence). In contrast, direct Sanger sequencing of the PCR product amplified from the clonal population yielded a sequencing chromatogram with overlapping peaks at the region of interest (top chromatogram). Analysis of this sequencing data with a commonly used bioinformatics tool indicated that different alleles could be present in the sample at various probabilities. Using the Guide-it Indel Identification Kit, it was determined that the clone had one correctly edited WT allele (WT silent; middle chromatogram) and one indel allele with a one-base insertion (G) together with a 7-bp deletion (bottom chromatogram), a result consistent with the output of the Guide-it Knockin Screening Kit.
Beta-tester data
Comparing results of the knockin screening kit with Sanger sequencing in clonal cell lines
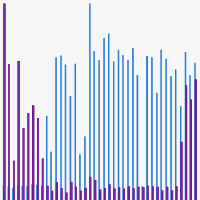
Dr. Justin McDonough, from the Jackson Laboratory, compared the performance of the Guide-it Knockin Screening Kit vs. software-based analysis of Sanger sequencing data obtained from PCR products for the detection of heterozygous (SNP/WT) edited iPSC clones (Figure 6). When interpreting these results, it must be taken into account that the detection of only one fluorescent signal (green or red) in a clonal cell line is correlated to the presence of at least one copy of the corresponding allele and indicates the absence of the other allele being assayed. For example, the genotype of a clone that generates only a green signal is SNP/SNP or SNP/Indel but not WT/SNP, WT/WT, or WT/Indel.
While the results of the Guide-it Knockin Screening Kit were consistent with the analysis of the Sanger sequencing traces for a majority of the clones (e.g., heterozygous clone A07), there were several instances where analysis of the Sanger data with a commonly used bioinformatics tool missed or called some clones incorrectly. For example, one clone carrying the SNP could not be identified by analysis of the Sanger data (F04; designated "unknown" in Figure 6). In addition, another clone carrying the desired SNP (E07) was called incorrectly to encode two copies of indel alleles at the target site. The beta tester indicated that, "the kit was very good at identifying WT/SNV hets, which is often what I'm looking to uncover."
Figure 6. Beta-tester data: successful identification of heterozygous edited clones. Genotypes determined via bioinformatic analysis of the Sanger sequencing data are indicated along the X-axis (WT, wild-type; SNP, accurate HDR; Indel, NHEJ; unknown, software could not determine). The presence of edited (SNP) and wild-type (WT) alleles in the clones as determined by the Guide-it Knockin Screening Kit are demonstrated by fluorescence signal intensities indicated in blue (green fluorescence) and purple (red fluorescence), respectively. While the results of the knockin screening were consistent with the bioinformatic analysis of the Sanger sequencing traces for a majority of clones analyzed, there were several instances where the Sanger sequencing analysis missed or called some clones incorrectly.
Conclusions
For researchers seeking to elucidate the genetic basis of diseases or develop novel therapies, the ability to engineer disease-associated SNPs using CRISPR-based genome editing technology in tandem with HDR has opened many doors. However, this approach is not without its challenges, due in part to the tendency towards NHEJ-based repair over HDR, and the difficulty associated with identifying successful HDR via Sanger sequencing of PCR products amplified from edited cells (even with the assistance of bioinformatic software tools). The Guide-it Knockin Screening Kit was developed with these challenges in mind. It provides researchers with a simple, rapid method to unequivocally identify successful HDR, both in bulk-edited populations prior to the tedious, time-consuming isolation of single-cell clones, and in the resulting clonal populations. As demonstrated by the data presented above, the knockin screening assay identifies successful HDR with greater certainty than direct Sanger sequencing and enables simultaneous detection of two different alleles (e.g., SNP and WT), a key benefit for researchers seeking to engineer cell lines that are heterozygous for a SNP allele of interest.
References
Kim, E. J., Kang, K. H. & Ju, J. H. CRISPR-Cas9: a promising tool for gene editing on induced pluripotent stem cells. Korean J. Intern. Med. 32, 42–61 (2017).
Paquet, D. et al. Efficient introduction of specific homozygous and heterozygous mutations using CRISPR/Cas9. Nature533, 125–129 (2016).