Modeling disease progression in an accurate and reproducible manner is a challenge for researchers studying diabetes and other metabolic disorders. Currently, the gold standard in diabetes modeling and drug discovery involves the use of primary pancreatic islets of Langerhans, small communities of hormone-producing cells that include insulin-producing beta cells. Aside from being difficult and costly to obtain, primary islets come from donors with different genetic backgrounds, confounding results by adding variability to cellular models and assays. As an alternative to islets, a rapidly renewable source of human induced pluripotent stem (hiPS) cell-derived beta cells from a single donor could be used as a powerful screening tool for drug discovery and as a physiologically relevant model of insulin production and release.
To address the need for a consistent and reliable source of beta cells, we have developed a standardized, four-step differentiation protocol with optimized culture conditions that mimics the typical stages of pancreatic development (Figure 1). hiPS cells are first differentiated into definitive endoderm cells. Following endodermal commitment, the cells are differentiated into pancreatic endoderm, and then further differentiated into endocrine progenitor cells. Finally, the progenitors undergo maturation into terminally differentiated beta cells.
Figure 1. hiPS cell-derived beta cell differentiation. Our differentiation strategy is based on a four-step protocol that resembles normal embryonic development of beta cells. hiPS cells are first differentiated into Sox17-positive definitive endoderm. Following endodermal commitment, the cells are further differentiated into Pdx1/Nkx6.1 double-positive pancreatic endoderm. Next, differentiation continues into endocrine progenitor cells, which are Ngn3/Nkx2.2 double-positive. Finally, the progenitors mature into terminally differentiated beta cells that are double positive for C-peptide and MafA. Abbreviations: Sox17 = SRY (sex determining region Y)-box 17; Pdx1 = pancreatic and duodenal homeobox 1; Nkx6.1 = NK6 homeobox 1; Ngn3 = Neurogenin 3; Nkx2.2 = NK2 homeobox 2; C-peptide = Connecting peptide; and MafA = MAF BZIP Transcription Factor A.
Two Cellartis hiPS beta cell kits are available, each containing iPS-derived beta cells from a different human donor line (ChiPSC12 and ChiPSC22; abbreviated C12 and C22, respectively). Each line is provided as a complete kit containing coating substrate, basal media, and a supplement.
Access to two lines of renewable, reliable beta cells allows functional studies that reflect individual variation and confidence in results across experiments. Beta cells from both donor lines also exhibit characteristics of mature beta cells and demonstrate low batch-to-batch variability. One variable of interest to diabetes researchers is the donor HLA type. Epidemiological studies have suggested that some common class I alleles, including HLA-A*02:01, may confer increased susceptibility to type 1 diabetes (Marron et al. 2002). The HLA types of C12 and C22 can be seen in Table I.
Cellartis hiPS Beta Cells (from ChiPSC12)
Cellartis hiPS Beta Cells (from ChiPSC22)
HLA-A*01:01
HLA-A*02:01
HLA-B*08:01, HLA-B*37:01
HLA-B*07:02, HLA-B*40:01
HLA-C*06:02, HLA-C*07:01
HLA-C*03:04, HLA-C*07:02
HLA-DRB1*03:01, HLA-DRB1*11:04
HLA-DRB1*13:02, HLA-DRB1*14:01
HLA-DQB1*02:01, HLA-DQB1*03:01
HLA-DQB1*05:03, HLA-DQB1*06:04
HLA-DPB1*01:01, HLA-DPB1*04:01
HLA-DPB1*03:01, HLA-DPB1*04:01
Table I. Donor information and HLA typification data for ChiPSC12 and ChiPSC22. The beta cells originate from donors with different HLA types. ChiPSC12 was sourced from skin fibroblasts from a healthy 24-year-old European/North African male volunteer (76 kg/177 cm). ChiPSC22 was sourced from skin fibroblasts from a healthy 32-year-old European/North African male volunteer (74 kg/179 cm) with a diabetes-susceptible HLA type, HLA-A*02:01.
In this technical note, we present data demonstrating the suitability of these hiPS cell-derived beta cells for drug discovery and for modeling of diabetes and other metabolic disorders.
We first assayed the hiPS cell-derived beta cells for the expression of mRNA transcripts known to be present in mature beta cells. All analyzed transcripts were present in the hiPS cell-derived beta cells, and the expression levels were compared to the expression levels in primary islets (Figure 2). Critically, all three batches of hiPS cell-derived beta cells from each donor cell line expressed mRNA at comparable levels, demonstrating high batch-to-batch reproducibility.
Figure 2. hiPS cell-derived beta cells consistently express key beta cell mRNAs. mRNA expression analysis, as quantified by RNA-seq, was performed on Cellartis beta cells from C12 and C22 on Day 14 post-thawing (n = 3 batches per cell line) and on primary islets (n = 5 donors). Abbreviations: PDX1 = pancreatic and duodenal homeobox 1; GCK = glucokinase; SLC2A1, SLC2A3 = Solute Carrier Family 2 Member 1 and 3; MAFB = MAF BZIP Transcription Factor B; NKX2.2 = NK2 homeobox 2; NKX6.1 = NK6 homeobox 1; and NEUROD1 = Neurogenic differentiation 1. Data are presented as the mean values ± SEM.
hiPS cell-derived beta cells synthesize insulin
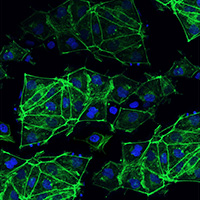
The main function of beta cells is to synthesize and secrete insulin. In the insulin synthesis pathway, proinsulin is cleaved into one C-peptide molecule and one insulin molecule (1:1 molar ratio) (Ashby & Frier 1981; Horwitz et al. 1975).Coexpression of C-peptide and insulin can, therefore, be considered an indication of functional insulin production. Here, we confirm that insulin is synthesized by the cells, as shown by the high correlation between insulin and C-peptide expression in beta cells derived from both C12 and C22 (Figure 3).
Figure 3. hiPS cell-derived beta cells synthesize insulin. Staining for C-peptide and insulin was performed by immunofluorescence on Cellartis beta cells at Day 14 post-thawing. Colocalization of insulin and C-peptide indicates intracellular insulin synthesis. Scale bar = 100 μm.
Several transcription factors play a critical role in beta cell development, proliferation, and functional activity. Developing and mature beta cells express Pdx1, Nkx6.1, and insulin. It has been suggested that Pdx1 acts as a master regulator of beta cell fate, activating genes essential for beta cell identity and repressing genes associated with alpha cell identity (Gao et al. 2014), whereas Nkx6.1-deficient cells acquired molecular characteristics of delta cells (Taylor, Liu, and Sander 2013).
As a marker for mature beta cells, MafA is distinct from other transcription factors since it is expressed exclusively in adult, mature beta cells and not during beta cell differentiation. Additionally, coexpression of insulin and Nkx6.1 is considered to be indicative of mature stem-cell-derived beta cells (Ma et al. 2018; Pagliuca et al. 2014; Rezania et al. 2014). Here, we show that Nkx6.1, Pdx1, and MafA, key markers of beta cells, are coexpressed with insulin and/or C-peptide in our hiPS cell-derived beta cells (Figure 4).
Figure 4. hiPS cell-derived beta cells coexpress key beta cell proteins. Staining for insulin and Nkx6.1 (Panel A), insulin and Pdx1 (Panel B), and C-peptide and MafA (Panel C) was performed on beta cells on Day 14 post-thawing. Scale bar = 100 μm.
We also quantified the number of cells coexpressing insulin and Nkx6.1 using flow cytometry (Figure 5). Notably, the percentage of insulin/Nkx6.1 double-positive cells are different in beta cells derived from C12 and C22, indicating variability between the two cell lines.
Figure 5. Quantitative determination of insulin/Nkx6.1 double-positive cells in hiPS cell-derived beta cells. Staining for insulin and Nkx6.1 was performed on beta cells from C12 or C22 on Day 14 post-thawing and analyzed by flow cytometry (FC). FC plots are shown for the analysis of one batch of beta cells from C22, with ~33% insulin/Nkx6.1 double-positive cells (Panel A) and ~56% insulin-positive cells (Panels A and B). Percentages of cells expressing insulin and coexpressing insulin/Nkx6.1 are shown for beta cells derived from C12 and C22, respectively (Panel C). Data are presented as mean values ± SEM (n = 3 batches from each cell line).
hiPS cell-derived beta cells secrete C-peptide
Pharmacological strategies for the treatment of type 2 diabetes include stimulation of insulin secretion. Glucose-stimulated insulin secretion (GSIS) relies on a functional potassium channel (KATP channel) and calcium channel (Ca2+ channel). Glucose enters the beta cell and causes ATP production via the TCA cycle in the mitochondria. The increased intracellular ATP/ADP ratio controls the closure of the KATP channel, leading to depolarization of the cell membrane and opening of the Ca2+ channel. The increased calcium ion concentration in the cell subsequently induces insulin secretion from storage vesicles (Figure 6).
Other potential druggable targets for diabetes are G protein-coupled receptors (GPCRs) expressed in islet cells (Ahrén 2009). In addition to glucose stimulation, a variety of mediators can induce insulin secretion; most of them act through GPCRs (Winzell and Ahrén 2007). Examples of islet GPCRs are glucagon-like peptide-1 (GLP1), the receptor for incretin hormones, the receptor for the classical neurotransmitter acetylcholine (AChR), and the GPCRs with fatty acids as ligands, i.e., FFAR1, FFAR2, and FFAR3 (Figure 6).
Figure 6. Schematic of beta cell insulin pathways. Several pathways regulate insulin synthesis and secretion in beta cells. Diagram was adapted from Yaluri et al. 2015, which can be accessed here, and is used under Creative Commons Attribution 4.0 International license.
Response to drugs targeting K+ and Ca2+ channels
To test beta cell functionality in response to a forced closure of the potassium channels, we exposed the cells to potassium chloride (KCl). Increased levels of KCl in the cell culture medium cause an increase in the intracellular level of potassium ions and the subsequent closure of the KATP channel. We also evaluated the combinatory effect of the KCl and isradipine, a Ca2+ channel blocker that counteracts the effect of KCl (Figure 7). Here, and in all following experiments, C-peptide secretion into the culture media was measured by ELISA and used as an indicator of insulin secretion.
Figure 7. hiPS cell-derived beta cells show a physiologically relevant response to drugs targeting the potassium and calcium channels. hiPS cell-derived beta cells from C12 or C22 were incubated in 2.8 mM glucose (control), and then exposed to KCl alone or KCl and isradipine. Amounts of C-peptide detected in the culture media were normalized to the matched control. Data are expressed as mean ± SEM (n = 6); *** = p<0.001.
Response to drugs that stimulate GPCRs
To further evaluate the functionality of our hiPS cell-derived mature beta cells, we tested their response to two drugs that stimulate different GPCRs. We incubated the cells with exenatide (an incretin mimetic) or acetylcholine, which are both known to increase insulin secretion in islets (Layden, Durai, and Lowe, Jr. 2010). Exenatide is used in the treatment of diabetes mellitus type 2 and binds to the GLP-1 receptor, and acetylcholine binds to the acetylcholine receptor. The cells responded to stimulation by exenatide (Figure 8, Panel A) and acetylcholine (Figure 8, Panel B) with increased C-peptide secretion.
Figure 8. hiPS cell-derived beta cells secrete C-peptide in response to exenatide and acetylcholine.Panel A. Beta cells from C12 or C22 were incubated in 5.5 mM glucose. Addition of 100 nM exenatide for 45 min resulted in an increase in C-peptide secretion. Panel B. Beta cells from C12 and C22 were incubated in 5.5 mM glucose. Addition of 25 μM acetylcholine for 45 min resulted in an increase in C-peptide secretion. Treatment levels were normalized to the matched control. Data are presented as mean values ± SEM (n = 4); * = p<0.05, ** = p<0.01.
Next, we investigated beta cell functionality in response to short-chain fatty acids, which act as ligands to GPCRs. In islets, FFAR2 (GPR43) and FFAR3 (GPR41) are the two GPCRs that bind short-chain fatty acids. The relationship between FFAR2/3 agonism and insulin secretion is not fully understood, but studies report an increase in insulin secretion upon activation of FFAR2 (Bolognini et al. 2016; Priyadarshini et al. 2015; Veprik et al. 2016). To evaluate the effect of FFAR2 agonists, we used the allosteric FFAR2 modulator, 4-CMTB. Figure 9 shows that C-peptide secretion increased in response to 4-CMTB, indicating that the hiPS cell-derived beta cells respond to the stimulus in the same manner as has been observed in primary islet cells.
Figure 9. hiPS cell-derived beta cells secrete C-peptide in response to FFAR2 agonist stimulation. hiPS cell-derived beta cells from C12 or C22 were incubated in 5.5 mM glucose. Addition of 150 μM 4-CMTB for 45 min resulted in an increase in C-peptide secretion. Treatment levels were normalized to the matched control. Data are expressed as mean values ± SEM (n = 6); * = p<0.05, *** = p<0.001.
Conclusions
To date, human primary islets have been considered the gold standard for diabetes research, as they provide a model to assess key criteria such as insulin production and insulin secretion. However, availability of these cells can be limiting. Moreover, primary islets can be unreliable: they show large donor variation, preventing the generation of consistent results. Beta cells derived from human induced pluripotent stem cells are a powerful alternative to primary islets, as they offer a virtually unlimited source of cells, provided they recapitulate key beta cell functions.
Cellartis hiPSC-derived beta cells are available from two different hiPS cell lines from healthy donors (C12 and C22). The cells express insulin, C-peptide, MafA, Nkx6.1, and Pdx1; have functional potassium and calcium channels; and respond to multiple GPCR agonists including exenatide and acetylcholine. Combined with the fact that they are easy to culture, Cellartis beta cells are an excellent tool for modeling diabetes and other metabolic disorders, as well as for supporting screening efforts for drug discovery.
Total RNA was extracted from Cellartis beta cells from C12 and C22 (n = 3 batches per cell line) as well as from primary islets (n = 5 donors) using the GenElute RNA/DNA/Protein Plus Purification Kit (Sigma Aldrich). Samples were sequenced on an Illumina HiSeq® 2500 using the HiSeq SBS Kit v4. Reads were mapped to the human reference genome GRCh37, and read counts were obtained using the featureCounts program in the R subread package (Liao, Smyth, and Shi 2013). Read counts were normalized to FPKM values by StringTie (Pertea et al. 2015).
Immunocytochemistry
Cells were fixed in 4% paraformaldehyde, permeabilized in 0.3% Triton X-100, blocked in TNB blocking buffer (PerkinElmer), and immunolabelled with primary antibodies specific to the marker of interest: insulin ab (Abcam), C-peptide ab (Cedarlane Laboratories), MAFA ab (Abcam), and/or PDX1 ab (Abcam). All primary antibodies were diluted 1:500 in TNB blocking buffer except for PDX1, which used a 1:8,000 dilution. Fluorescent-conjugated secondary antibodies (Alexa Fluor 488 or Alexa Fluor 594, Thermo Fisher Scientific) specific to the primary antibody host species were added to visualize the marker of interest. The secondary antibody was diluted 1:1,000 in TNB blocking buffer.
KCl/isradipine response
In Stage 1, cells were incubated in KREBS buffer with 2.8 mM glucose for 45 min (control). In Stage 2, the same buffer was supplemented with 30 mM KCl and the cells were incubated for 45 min. In Stage 3, both KCl and isradipine (10 μM) were added, and the cells were incubated for 45 min. The samples were collected at the end of each stage, and C-peptide secretion was measured using the Mercodia C-peptide ELISA (which recognizes human C-peptide). The experiment was performed on two batches of Cellartis beta cells and three biological samples per batch.
Flow cytometry
Cells were dissociated, fixed in 4% paraformaldehyde, and stored overnight in 90% MeOH at –20°C. For the staining, cells were washed and probed with Biotin-conjugated Insulin mAb (Abcam), diluted 1:400, and incubated with Alexa Fluor 488-conjugated NKX6-1 ab (Novus Biologicals), diluted 1:10,000. After repeated washing, PE-conjugated Streptavidin (Abcam), diluted 1:4,000, was used to visualize the biotin-conjugated Insulin mAb. The flow cytometry analysis was performed on a Guava easyCyte flow cytometer (Millipore Sigma) and analyzed using the GuavaSoft Software Package (Millipore).
Exenatide, acetylcholine, and 4-CMTB stimulation
In Stage 1, cells were incubated in KREBS buffer with 5.5 mM glucose for 45 min (control). In Stage 2, the same buffer was supplemented with exenatide (100 nM), acetylcholine (25 µM), or 4-CMTB (150 µM), and then the cells were incubated for 45 min. Samples were collected at the end of each stage, and C-peptide secretion was measured by Mercodia C-peptide ELISA.
References
Ahrén, B. Islet G protein-coupled receptors as potential targets for treatment of type 2 diabetes. Nat. Rev. Drug Discov.8, 369–385 (2009).
Ashby, J. P. & Frier, B. M. Circulating C-peptide: measurement and clinical application. In Ann. Clin. Biochem (Vol. 18) (1981).
Bolognini, D. et al. The Pharmacology and Function of Receptors for Short-Chain Fatty Acids. Mol. Pharmacol. 89, 388–398 (2016).
Gao, T. et al. Pdx1 Maintains β Cell Identity and Function by Repressing an α Cell Program. Cell Metab. 19, 259–271 (2014).
Horwitz, D. L. et al. Proinsulin, insulin, and C-peptide concentrations in human portal and peripheral blood. J. Clin. Invest.55, 1278–1283 (1975).
Layden, B. T., Durai, V. & Lowe, W. L. G-Protein-Coupled Receptors, Pancreatic Islets, and Diabetes. Nat. Educ. 3, 13 (2010).
Liao, Y., Smyth, G. K. & Shi, W. The Subread aligner: fast, accurate and scalable read mapping by seed-and-vote. Nucleic Acids Res. 41, e108-e108 (2013).
Ma, S. et al. β Cell Replacement after Gene Editing of a Neonatal Diabetes-Causing Mutation at the Insulin Locus. Stem Cell Rep.11, 1407–1415 (2018)
Marron, M. P. et al. Functional evidence for the mediation of diabetogenic T cell responses by HLA-A2.1 MHC class I molecules through transgenic expression in NOD mice. Proc. Natl. Acad. Sci. U. S. A. 99, 13753–8 (2002).
Pagliuca, F. W. et al. Generation of Functional Human Pancreatic β Cells In Vitro. Cell159, 428–439 (2014).
Pertea, M. et al. StringTie enables improved reconstruction of a transcriptome from RNA-seq reads. Nat. Biotechnol. 33, 290–295 (2015).
Priyadarshini, M. et al. An Acetate-Specific GPCR, FFAR2, Regulates Insulin Secretion. Mol. Endocrinol. 29, 1055–66 (2015).
Rezania, A. et al. Reversal of diabetes with insulin-producing cells derived in vitro from human pluripotent stem cells. Nat. Biotechnol.32, 1121–1133 (2014).
Taylor, B. L., Liu, F. F. & Sander, M. Nkx6.1 is essential for maintaining the functional state of pancreatic beta cells. Cell Rep. 4, 1262–75 (2013).
Veprik, A. et al. GPR41 modulates insulin secretion and gene expression in pancreatic β-cells and modifies metabolic homeostasis in fed and fasting states. FASEB J. 30, 3860–3869 (2016).
Yaluri, N. et al. Simvastatin Impairs Insulin Secretion by Multiple Mechanisms in MIN6 Cells. PLoS One10, e0142902 (2015).