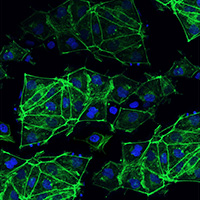
Robust expansion of clones The Cellartis DEF-CS 500 Culture System promotes a high survival rate of iPS cell colonies and robust proliferation of iPS cell clones.
Induced pluripotent stem (iPS) cells are generated by reprogramming a differentiated somatic cell into a pluripotent stem cell. This source of pluripotent cells enables drug discovery, disease modeling, and ultimately gives us a chance to find cures for today’s most devastating diseases. With iPS cells, researchers can generate patient- or disease-specific cells, enabling new frontiers in disease modeling (Meissner, Wernig, and Jaenisch 2007).
Although there are several methods for reprogramming cells, the Sendai-virus method was shown to be highly reliable and efficient in comparison to other non-integrating methods (Schlaeger et al. 2015). The most commonly used reprogramming factors are the Yamanaka factors, Oct3/4, Sox2, c-Myc, and Klf4, which are suitable for efficient reprogramming (Takahashi et al. 2007).
The Cellartis DEF-CS 500 Culture System promotes reliable growth of hiPS cells in a defined, feeder-free environment. Cells are grown as a homogeneous monolayer and are enzymatically passaged as single cells that maintain pluripotency with a stable karyotype for more than 20 passages (Asplund et al. 2016). Here we show that the Cellartis DEF-CS 500 Culture System efficiently and robustly reprograms human primary peripheral blood mononuclear cells (PBMCs) using Sendai virus in a 21-day protocol (Figure 1).
Figure 1. Reprogramming timeline with the Cellartis DEF-CS 500 Culture System.
Results
Using our PBMC reprogramming protocol (Figure 1), we reprogrammed seven different human PBMC cultures using Sendai virus and the Cellartis DEF-CS 500 Culture System. First, human PBMCs from ATCC were reprogrammed to evaluate the method, then PBMCs from 6 different donors (A–F) were reprogrammed. Most of the data shown are from A–F, which were analyzed in greater detail.
For all tested PBMCs, stable iPS cell lines were generated on the first transduction attempt. We transduced on the order of 106 PBMC cells from each donor, which generated large numbers of colonies, and picked 8–30 colonies from each of the donors for further culturing.
The survival rate of picked colonies from Donors A–F varies between 40–75%, depending on the donor, according to Table I. Here, the survival rate reflects the number of healthy, pluripotent colonies with robust growth.
Table I. The survival rate of picked colonies from donors A–F.
Survival rate of reprogrammed colonies
Donor
A
B
C
D
E
F
Picked colonies
15
12
12
12
8
8
Generated iPS cell lines
6
9
8
5
6
6
Generated iPS cell lines (%)
40
75
67
41
75
75
The morphology of the cells was monitored regularly. Figure 2 shows the morphological changes of a colony from the day of picking until the clone is ready to be frozen as a seed bank at Passage 6.
Figure 2. The progress from a colony at the day of picking to a clone ready to be frozen as a seed bank.
One generated iPS cell line from each donor—six iPS cell lines in total—were further passaged to study the loss of the Sendai virus. At some passages the amount of Sendai virus RNA was measured in the cells using quantitative real-time PCR. Ct values ≥36 were considered negative. At Passage 7–8, two-thirds of the iPS cell lines were cleared from Sendai virus RNA. All tested iPS cell lines were cleared from Sendai virus RNA by Passage 18 (see Figure 3).
Figure 3. Sendai virus is undetectable after Passage 18. Sendai virus RNA in the iPS cell lines was continuously measured using quantitative real-time PCR. Ct values ≥36 were considered negative.
At Passage 15, the iPS cell lines were analyzed for the stem cell markers Oct4, SSEA-4, TRA-1-60, and TRA-1-81, using immunocytochemistry (ICC). A vast majority of the cells were positive for all four markers (see Figure 4), whereas no expression of SSEA-1 was observed.
Figure 4. Immunocytochemical analysis for the detection of the stem cell markers Oct4, SSEA-4, TRA-1-60 and TRA-1-81 in one of the iPS cell lines at Passage 15. SSEA-1 was used as a negative marker. DAPI was used for nuclear staining. Overlap of DAPI and markers is shown in purple.
At Passages 14–17, the six iPS cell lines were also analyzed for the expression of Oct4 by flow cytometry. The cells contained a high proportion of Oct4-positive cells, according to Figure 5.
Figure 5. The 6 iPS cell lines were analyzed for the expression of Oct4 by flow cytometry. All cell lines were nearly 100% positive for Oct4. Panel A. Representative FACS plot. Panel B. The percentage of cells positive for Oct4 varied in a narrow range between 99.58% and 99.94%.
To further examine the pluripotency of the six generated iPS cell lines, they were differentiated into Definitive Endoderm (DE) cells and then further into hepatocytes. All iPS cells generated DE cells and hepatocytes. ICC was performed to confirm differentiation to DE using the DE marker SOX17 and pluripotency marker Oct4. The majority of the cells in each iPS line were differentiated to DE cells, as indicated by positive staining for SOX17 and negative for Oct4 at Day 7, shown in Figure 6.
Figure 6. Differentiation to DE indicated by morphology, the DE marker Sox17 (red), and the pluripotency marker Oct4 (green). The majority of cells from each iPSC line was differentiated to DE cells. DAPI was used for nuclear staining.
ICC was also performed to confirm differentiation to hepatocytes after 30 days of differentiation, as indicated by the hepatic markers HNF4α, albumin, and CYP3A4. The majority of the cells stained positive for HNF4α. A subset of the hepatocytes was also positive for albumin or CYP3A4, with only a few double-stained cells (see Figure 7).
Figure 7. Differentiation to hepatocytes after 30 days of differentiation indicated by morphology, the hepatic markers HNF4α, albumin (green), and CYP3A4 (red). The majority of the cells stained positive for HNF4α. A subset of the hepatocytes was also positive for albumin or CYP3A4, with only a few double-stained cells. DAPI was used for nuclear staining.
Conclusion
Using the Cellartis DEF-CS 500 Culture System when reprogramming human PBMCs with Sendai virus allowed for generation of multiple iPS cell lines from each donor with only one transduction step. The Cellartis DEF-CS 500 Culture System promoted high rates of survival in the emerging iPS cell colonies and robust proliferation into iPS cell lines. The reprogrammed iPS cell lines showed high expression levels of the stem cell markers Oct4, SSEA-4, TRA-1-60, and TRA-1-81, and the typical morphology of undifferentiated stem cells. In addition, the generated iPS cells were shown to differentiate into definitive endoderm and hepatocytes when exposed to the Cellartis Definitive Endoderm Differentiation Kit with DEF-CS Culture System and the Cellartis Hepatocyte Differentiation Kit. The highly efficient and reliable performance of the Cellartis DEF-CS 500 Culture System in these studies demonstrates the time- and cost-effectiveness of this method.
Methods
Culture of PBMCs
The PBMCs were thawed and seeded in PBMC medium. They were maintained in PBMC medium for four days prior to the start of the Sendai reprogramming.
Sendai reprogramming and transfer to the Cellartis DEF-CS 500 Culture System
The CytoTune-iPS 2.0 Sendai Reprogramming Kit (Fisher Scientific) was used to reprogram the PBMCs. The reprogramming vectors include the four Yamanaka factors, Oct3/4, Sox2, c-Myc, and Klf4. The major reprogramming steps are shown in Figure 1. Plating of the PBMCs, transduction, and maintenance of transduced cells until Day 3 were performed according to the kit user guide. At Day 3 post-transduction, the transduced cells were plated on COAT-1-coated culture dishes (dilution 1:5) in PBMC medium. During Days 7–8, the medium was changed to the complete DEF-CS medium. This medium was replaced daily until the colonies were ready to be picked.
Picking and expansion of colonies
Three to four weeks post transduction the colonies were ready to be manually cut and transferred to fresh COAT-1-coated culture dishes. The clones were then cultured according to the Cellartis DEF-CS 500 Culture System User Manual.
Quantitative real-time PCR
Gene expression was analyzed using the TaqMan Gene Expression Assay SEV (Sendai virus) and the amplification reactions were carried out in a real-time PCR system. The samples were normalized to the expression of CREBBP (CREB binding protein).
Immunocytochemistry
The iPS cell lines were stained with Oct4, SSEA-4, TRA-1-60, TRA-1-81, and SSEA-1 antibodies. The DE cells were stained with SOX17 and Oct4 antibodies. The hepatocytes were stained with HNF4α, albumin, and CYP3A4 antibodies. The cells were further stained with secondary antibodies conjugated with Alexa Fluor 488 or 594 and DAPI. Staining was examined using a fluorescence microscope.
Flow cytometry
The iPS cell lines were fixed in 4% formaldehyde and then stained with Oct4 antibody and a secondary antibody conjugated with Alexa Flour 488. The labeled cells were analyzed using flow cytometry on a Guava EasyCyte 5HT, and the percentage of positive cells was quantified with GuavaSoft Software.
Differentiation of iPS cells to definitive endoderm cells and hepatocytes
The differentiation of the iPSC cell lines to DE cells and hepatocytes were performed using the Cellartis Definitive Endoderm Differentiation Kit with the DEF-CS Culture System (Cat. No. Y30035) and the Cellartis Hepatocyte Differentiation Kit (Cat. No. Y30050) according to their respective user manuals.
References
Asplund, A. et al. One Standardized Differentiation Procedure Robustly Generates Homogenous Hepatocyte Cultures Displaying Metabolic Diversity from a Large Panel of Human Pluripotent Stem Cells. Stem Cell Rev. Reports12, 90–104 (2016).
Meissner, A., Wernig, M. & Jaenisch, R. Direct reprogramming of genetically unmodified fibroblasts into pluripotent stem cells. Nat. Biotechnol.25, 1177–81 (2007).
Schlaeger, T. M. et al. A comparison of non-integrating reprogramming methods. Nat. Biotechnol.33, 58–63 (2015).
Takahashi, K. et al. Induction of Pluripotent Stem Cells from Adult Human Fibroblasts by Defined Factors. Cell131, 861–872 (2007).